- 程序有所改变。发帖如还有问题请报告
- 【征集】西西河的经济学,及清流措施,需要主动参与者,『稷下学宫』新认证方式,24年网站打算和努力目标
主题:261-Philip Ball:水为什么这么怪? -- 万年看客
https://www.youtube.com/watch?v=2IlucL05bhc&list=PL4i9YSoIJiPfAq5TCk7xdVrJlxRAMbay-&index=109&t=3229s
我在《自然》杂志工作了很久,1988年就加入了杂志社。当时我刚刚写完了关于液态物理学的博士论文,但是我没想到这篇论文与我在《自然》杂志社将要从事的工作有什么关系。但是我刚刚开始工作,事实就证明了我的错误。因为在此之前几周,《自然》发表一篇论文——现在这篇文章已经臭名昭著了——声称水有记忆。论文的作者是法国免疫学家杰克斯.宾文尼斯特(Jacques Benveniste)和同事们,他们声称在高度稀释的抗体溶液当中发现了生物行为,尽管这些溶液被稀释到了根本就不会包含抗体分子的地步。更奇怪的是,这种行为看起来有时会出现,有时会消失,似乎会随着稀释而不断涨落。乍一看去这个现象根据已知的化学原则没有任何可行的解释,没有分子就应该没有效果。宾文尼斯特认为自己有个解释:“通过无限的氢键网络或者电磁场,水可以成为其他分子的模板。”这话啥意思?我马上就跟大家解释他们以为这段话是什么意思,不过当时我认为这是《自然》刊登过的最没有意义的句子。或者说,这句话的意义并不在词语当中,而是藏在词语背后。宾文尼斯特等人设法援引了液态水的奇怪化学特质,听上去足够科学,以至于瞒过了审稿人的雷达——我很怀疑审稿人并不非常了解液态理论。他们利用了水的无可争议的怪异特质。
宾文尼斯特绝不是第一个或者最后一个这么做的人,我只是拿这个故事举个例子而已。你们肯定能想到,宾文尼斯特的论文很快就为所谓顺势疗法提供了支持。顺势疗法主张将药物大量稀释,直到溶液当中几乎不含有效成分,但是这样的溶液依然可以用来治病。因此这篇论文不仅争议缠身,而且还遭到了政治化对待。《自然》刊发这片论文之后,编辑约翰.麦多克斯(John Maddox)率领一支队伍造访了宾文尼斯特的试验室,观察了他们复制试验结果的尝试。结果宾文尼斯特的研究人员无法重现所谓的“生物特质随着溶液稀释而不断上升下降”现象。麦多克斯的调查团队因此认为宾文尼斯特之前的发现只是幻觉。宾文尼斯特则声称自己遭到了猎巫迫害。他坚持研究高度稀释溶液,终于提出了所谓的数字生物学,声称自己可以使用电磁信号通过电话发送生物行为,然后用这些信号为纯水编程,使得纯水成为生物信息的载体。他直到2004年去世前都在从事这方面的工作。今天依然有人坚称水有记忆,可以让高稀释度溶液具备原本不该有的性质。就连诺贝尔奖得主当中也有一位免疫学家吕克.蒙塔尼(Luc Montagnier)相信这一点。
关于水的特质的奇特主张,以上并不是孤立个案。这方面最著名的事件是聚合水丑闻。二十世纪六十年代,苏联化学家声称他们在玻璃细管里发现了一种新型的水,质地就像软蜡一样。他们甚至认为这可能是在日常温度与压力下最稳定的水的形态。他们提出理论,只要将一滴聚合水滴入大海,整片大海都会变成胶质,就像库特.冯内古特的小说《猫的摇篮》当中的虚构物质九号冰,足以冻住全世界的海水。但是事实证明,聚合水只是另一个幻象。所谓的蜡样物质只是浓缩的杂质,很可能是玻璃渗出的硅。凑巧的是,冯内古特笔下的九号冰是由一位费利克斯.霍尼克尔发明的,这个角色的原型显然是美国科学家欧文.朗缪尔。朗缪尔当年创造了一个词,用来形容聚合水与水有记忆之类的主张:他将这些主张称为“病态科学”。
水具有吸引病态科学的强大能力。有时这些病态科学会渗入科学文献的页面,有时则游走在科学边缘与神秘主义你中有我的边缘地带。这方面有一个非同寻常的故事,主人公是一位奥地利护林员,名叫维克多.肖伯格(Viktor Schauberger)。他认为旋涡的能量能让水获得生命。据说纳粹对于他的理念很感兴趣,要求他开发某种秘密武器,因为他认为漩涡会导致水的内爆。当前确实是新时代神秘主义与科技现实相互融合的时代。我听过其他科学家一口咬定特殊处理过的金属杖可以改变水的味道与粘稠度。我还发现,只要向非科学从业者提到我正在研究水的性质,用不了多久就会有人提到江本胜的《水知道答案》。他声称自己依靠祈祷改变了雪花的形状,并且写了一本畅销书来解释这一点。书中收录了一批从日本藤原大坝提取的水的冰冻样本。江本胜形容这批样本“黑暗,无形,无法结晶。”然后他请来高僧对着这批样本念了一个小时的佛经,新的样本就产生了十分规整的六角晶体。显然音乐也有影响:江本胜在水结晶时播放猫王的《伤心旅馆》,结果据他所说晶体自发地裂开了。
到底发生了什么?为什么水会吸引这么多怪人?科学家肯定能理解我下面的说法:研究水与研究其他物质并不一样。水并不是中立的物质。我们在研究水时全都携带着丰富的的文化联系,根本不可能抛在一旁。水确实是一种元素,不是化学元素,而是神话与诗歌当中的文化元素,而这点很重要。我推荐法国哲学家加斯东.巴什拉的《水与梦》,探索了各种水的诗歌化身在神话传说当中的深切根源。我相信,将水视作净化与救赎之力的理念构成了许多病态水科学背后的无意识心理。但是水之所以倾向于引发病态科学,并不仅仅因为它在文化当中占据中心地位,还因为水的性质确实很奇怪。我所指的是水与其他液体不一样,至少不像液态物理学家通常研究的大部分液体。
外人可能觉得很奇怪,不过我的博士论文的绝大部分篇幅都在讨论液体的氩或者氮,这两种液体绝大多数人从来都见不到,在常温常压环境下也根本不存在。但是这些液体很容易理解,也很容易针对他们进行建模。在液态的氩或者氮当中,分子或者原子就好比小球,彼此之间微弱地相互吸引。水与其他类似化合物相比则显然不同。水是氢化的氧,一个氧原子与两个氢原子结合。如果将水与元素周期表上氧元素附近其他元素的氢化物质相比——比方说甲烷、氨、氢氟酸、硫化氢以及盐酸——所有这些化合物在室温下都是气体,盐酸略微稳定一点,但是到了夏天高温的时候也会变成气体。但是水在室温下的沸点却高得多。此外水会结冰,冰会浮在水面。这一幕看上去如此司空见惯,以至于我们很容易忘记,其他所有液体一旦降温凝固密度都会增加,但是水在冰冻时却会膨胀,所以冬天的自来水管才会涨裂,所以泰坦尼克号才会沉没。更奇怪的是,冰并不会突然膨胀。水在冰冻之前就会略微膨胀一点点。一旦温度降到4摄氏度以下,水就会从热胀冷缩变成冷胀热缩。4摄氏度的水的密度最大,然后就开始降低,温度达到冰点时水的密度更是会陡然下降。因此冬天的湖底与上层相比要稍微暖和一点,也因为如此湖面才会从上往下冰冻,使得足够深的湖水不至于一冻到底,因为水面冰层反而起到了为水底保温的作用。
水的古怪性质可以列成一张清单,上面说的只是一般人最容易理解的特质。这一系列特质都可以通过一个原因来解释,正是这一点区分了水与液态氩之类的简单液体:水分子有某种粘稠性,因为分子之间可以形成微弱的化学链,也就是氢键。关于氢键的传统解释认为,氧原子倾向于将氢原子的电子云吸引向自己,使其与氢原子结合,使得氢原子略微带一点正电。而且一个氧原子有两个电子对不会参与形成化学键,因此会从氢氧结合一侧的背面探出头来,使得这一侧带负电。带正电的氢离子受到这两对电子的静电吸引,于是水分子之间就出现了氢键。这个解释其实过于简单,实际上我们至今都没能彻头彻尾地描述氢键。国际纯粹与应用化学联合会正在研究全新的氢键定义,企图将我们近年来通过最新研究获得的知识整合进新定义当中。但是旧定义作为粗略模型眼下也姑且够用了。
氢键并非水的专利,其他好几种简单氢化物都会形成氢键,包括氨与氢氟酸,它们也有略高于其他氢化物的沸点。但是所有水分子都可以形成至少四个氢键:两个氢原子各自形成一个,氧原子通过两个电子对形成两个,而且这四个氢键在空间当中各自指向特定的方向。不妨将水分子想象成一个小人,两只手——两个氢原子——各自抓住另一个小人的脚腕——氧原子的两个电子对之一,同时自己的两个脚腕也被另外两个小人抓住,于是摆出一个“大”字型。无数小人勾连在一起,就形成了网络结构,每一个水分子都会与周围其他水分子构成四面体结构,这个三维结构在空间里不断重复,形成了极大的网络。
由于氢键网络的存在,水在分子结构层面十分紧张。一方面,液体分子通常倾向于相互接近;另一方面,水分子又需要相互隔离一段距离才能形成氢键。这些氢键可以变形一点点,但是如果变形幅度太大就会破裂。水的怪异特性的根源就在于水分子无序紧密堆叠与有序四面体结构氢键网络之间的微妙平衡。实际上氢键网络的结构非常刚性,所以冰或者说水的结晶才会具备高度有序的结构。此外冰晶结构当中存在大量空隙,所以水结冰后才会膨胀。氢键网络同样解释了冰为什么有这么多不同的结晶形态——目前已经发现了十二种——因为在不同温度下,冰晶结构会受到不同程度的挤压,使得水分子可以通过各种方式堆叠起来,尤其可以通过在空隙之间形成第二套氢键网络更加紧密地堆叠起来。画面这张图片体现了这一点。想象一个舞池,里边有十八个小人,手拉手组成三个六边环,正好将舞池占满。假设我们在舞池里硬塞进两排椅子,舞池里依然可以站开十八个人,只要让一个六边环与另外两个重叠一部分。同理,高压条件下结成的冰也具备不同于常压冰的结构。所以就像库尔特.冯内古特在《猫的摇篮》当中描写的那样,确实存在九号冰,还存在十号冰和十一号冰。只不过现实生活当中的九号冰并不像冯内古特描写的那样邪性。某些形态的冰确实在0摄氏度以上依然是固体,但前提是必须处于高压环境下。
当冰融化时,有些氢键会断裂,分子相互靠近,致使水的密度升高。在液态水当中,氢键很不完美、很无序而且非常动态,不断出现与消失,每个氢键平均只能存在一皮秒,也就是一万亿分之一秒。研究水的化学家与物理学家一直很难恰如其分地形容液态水的氢键网络,因为这套网络完美地融合了秩序与非秩序。每一个水分子都大差不差地以四面体形态与周围的水分子协调一致。但是由于氢键很容易断裂,平均来说每个水分子都只能与少于四个的其他水分子相互联结。在冰晶当中水分子会形成六边环,但是在液态水当中最常见的却是五边环。还有些水分子能生成五个氢键,或者说形成分叉氢键,也就是让一个氢原子与两个氧离子连在一起。我要指出,即便这种常见的液态水的结构也并非完全确定。有科学家最近主张,液态水的分子可以连接成长链结构而不是三维网络,或者一部分水分子会组成比起其他分子更加致密的分子团,还有人主张液态水的分子结构是一块块冰晶状有序结构与大片无序结构的混合。根据我们目前掌握的知识,这些主张全都不太可能为真。但是这些主张的存在本身就意味着我们很难切断水的结构究竟是什么。
事实上,在高压低温的条件下确实很有可能存在两种液态水。此时液态不再是水的最稳定形态,很容易结冰。这是一个非常难以用实验来验证的理念,因为很难让水达到这种条件而不结冰。根据计算机模型,假如让水在这种条件下保持液态,那么水就会自动分离成为高密度形态与低密度形态,两者就像一般环境当中的水与油一样界限分明。这种有序结构与无序结构之间的紧张关系使得水转化成了两种不同的液体。水在4摄氏度时密度最大这一事实或许正是呼应了这两种形态。
正是因为水的液体形态如此复杂且充满不确定性,所以人们才会这么容易就经不住诱惑,采用五花八门的奇谈怪论来解释水的结构,杰克斯.宾文尼斯特恰恰正是想要这么做。因为水分子会形成三维网络,所以他才会暗示或许这套网络可以围绕一个有机分子“冻结”起来,“记住”这个分子的结构,将有机分子的形状“压印”在水分子网络上,而且有机分子的生物功能正是由形状决定的。可悲的是,这个理念毫无意义,因为完全没有理由认为印模的功能与造成印模的物体一致。毕竟,印模不是原件的复制品。话又说回来,水分子网络高度动态,氢键在一皮秒之间频繁出现而又消失,有机分子就算真能在网络上造成印模也必然无法持久。尽管如此,这样那样的水分子结构依然经常被人们拿来充当机械降神,用来解释各种异乎寻常的物理与生物学现象——通常都是在走投无路的情况下才这么办。
这种理念在生物学领域尤其臭名昭著,尤其是讨论水在细胞内部怎样表现的时候。直到今天这依然是个争议极大的领域。一方面,许多生化课本认为水只是生命的背景,他们承认水很重要,例如蛋白质分子没有水就维持不了结构与弹性,但是又认为水只是普适溶剂,解释完了三维氢键网络之后就不再多说什么。生物分子往往被描述成漂浮在虚空当中。为了清晰研究生物分子,这种呈现方式确实有道理,但是往往让人们以为无论这些分子周围有没有水分子,看上去都是这个样子。根据这种观点,细胞至多被视作装满被稀释物质的口袋。另一方面,也有人认为细胞里的水与一杯水里的水具有根本性的不同,而且这种不同对于细胞的健康至关重要。他们主张细胞里的水更粘稠。细胞里的水越像普通的水,细胞就越病态,比如癌细胞里的水就比正常细胞里的水更像普通的水。这些人认为细胞驯化了普通的水,改变了水的结构,向它赋予了亲生物质的特性。这种理念由来已久。诺贝尔生物学奖得主阿尔伯特.纳扎波尔蒂.圣捷尔吉将水称作“生命的矩阵”,他在二十世纪五十年代就谈到了细胞里的水的结构。其他人也讨论过水如何在蛋白质表面形成分层结构
这两个理念——水在细胞内外的性质完全一致,或者细胞内部的水具有根本性的不同特质——似乎都不对,事实要更加复杂也更加有趣。细胞内部的水的结构与特质似乎会在较为较微妙的方面受到生物分子的影响,也会受到细胞质里其他化学物质的影响,例如盐。但是此类影响仅仅发生在局部,而且只能在短距离起作用。满足这两个条件之后,水似乎确实可以调整自己来适应周围的生物分子,生物分子似乎也可以适应水。正是这一微妙互动让生命得以成为现实。接下来我想请大家略微领略一下这种互动过程。
首先我们先来看看细胞内部。画面上是细胞内部的放大绘图,是一团细胞质,各种生物分子正在水里游来游去。可以看到,细胞质非常拥挤——实际情况更糟糕,因为画面上省略了一切小分子,例如糖。平均来说,细胞质里的蛋白质大分子的相互间距平均来说只有一纳米,也就是百万分之一米,或者说隔着三四层水分子。因此细胞里的水分子填满了生物分子之间的全部空隙。那么这种填充对水有什么效果?这个问题的答案一言难尽。关键问题在于贴近大分子表面的水分子是什么样?蛋白质表面的化学性质非常多样,有些表面覆盖着溶于水的化学基团,可以形成氢键或者具有电荷,例如盐。这些表面具有所谓的亲水性。另一部分表面的化学特质更接近脂肪或者油类,具有所谓的疏水性。画面上是一个典型的蛋白质分子,其表面的亲水与疏水区域分别用蓝色与红色标注,颜色越浓就越倾向于对应的特质,白色则意味着中性。如果水的分子结构确实会在这样的蛋白质分子的亲水与疏水表面遭到改变,那么这样的改变可以往外延伸多远?究竟会起到什么效果?人们对于这一点很有有争议。
有一点很明确:水会促使疏水表面相互聚合在一起,换句话说水中的大量油性物质会自发聚合成一团。这个现象在大尺度上也能见到,最近我们发现火蚁会利用这一特质。火蚁的体表具有疏水性,于是大量火蚁会在水面上包成一团,形成类似筏子的结构,让它们能够在水面漂浮,从而避免蚁群在洪水中被淹死。在蛋白质分子的尺度上,疏水表面在水中的相互聚合使得蛋白质聚集在一起,形成了复杂且具有功能性的聚合结构,也使得单独一个蛋白质分子得以维持三维结构,从而发挥身为酶的作用。蛋白质的本质其实就是折叠起来的氨基酸链条,链条上的疏水部分往往会被塞进蛋白质分子内部,不至于碰到水。
这一点很好理解,但是驱动疏水表面相互吸引的力量是什么?教科书的答案基于1945年提出的理念:贴近疏水表面的水分子会表现得更有秩序,更像冰,使得贴近疏水表面的水分子排列成为氢键损失最小化的结构。换句话说,疏水表面凝聚了一座小冰山。1959年,化学家沃尔特.考兹曼(Walter Kauzmann)显示了这个过程如何发生。假设有两个表面疏水的粒子,表面都覆盖着一层水分子。当两个粒子相互靠近,两者接触部分的水分子层就会重叠,然后一部分水分子会被排挤出去,从原本的有序变成无序,导致熵增,因此这个过程很容易进行。根据考兹曼的解释,疏水表面在水中的相互吸引是由熵增驱动的。
这个模式的问题在于没有足够证据证明他是正确的,倒是很有理由认为他是错误的。因为水似乎不太可能在疏水表面附近形成更有序的结构,倒是有可能形成更奇怪的结构,看上去并不像一层冰。更有可能的情况是,在疏水表面附近或许存在非常薄的一层空间,其中几乎没有水分子,就像裹着一层气体。因此对于聚合现象的最新解释认为,两个表面疏水、裹着一层气体的粒子凑在一起之后,两者间隙当中的水就会突然全部蒸发。这个现象被称作毛细蒸发,我们在原则上很熟悉这个现象:两个疏水表面之间的极少量水的沸点不同于大量的水,所以很容易蒸发,然后两个疏水表面就会被表面张力拉在一起。出于显而易见的原因,这个过程被称作去湿转化。根据理论预测,当两个疏水表面相隔一纳米的时候就会发生这种事情。另外蛋白质在折叠时,去湿过程也会让蛋白质链的疏水部分挤在一起直接在一起。计算机模拟了两块疏水表面平板相互接近,确实出现了去湿过程。
但是目前的研究表明,当蛋白质链的疏水表面挤在一起时,去湿转化其实是个非常罕见的现象。画面上这个小蛋白质名叫蜂毒肽,是蜜蜂毒液当中的蛋白质,由四个管状结构结合在一起形成筒状。这个蛋白质分子的形成过程当中似乎当真发生了去湿转化,但是这种蛋白质似乎是非常少见的反例。画面上的另一种蛋白质名叫BphC,确实由两个蛋白质分子的非常平的疏水表面贴在一起形成。不过并没有证据证明两个蛋白质分子会突然碰在一起,两者之间的水分子并不会突然蒸发,只会逐渐流失。因此这种吸引或许必须倚靠其他机制才得以发生。
除了能让蛋白质分子维持结构以外,水还是塑形剂,让蛋白质分子具有足够的弹性。如果蛋白质太干燥,或者被从水中取出并放入其他溶剂,要么这个蛋白质分子要么会崩溃,要么因为过于僵硬而发挥不了功能。蛋白质分子表面总会附着一层水分子,而且这些水分子往往还会附着于蛋白质分子表面的特定部位。画面上的两张图片标注了水分子在不同蛋白质表面的分布,通过分析蛋白质分子晶体得以确定。水分子用黄色,绿色与紫色小球来表示,不同颜色代表了水分子位于蛋白质分子表面的不同层次。这种受束缚的水分子可以被视为生物分子结构的固有组成部分,这些水分子就像工具塑造了蛋白质的结构,同时其本身也具有很多特化功能。一方面,水分子是可以被取出的填充物,塞在蛋白质的空腔里,并且可以与其他分子结合,从而让蛋白质分子得以生冷不忌地与更多分子结合。比方说OppA蛋白质就可以借助水分子抓住许多二到五个氨基酸构成的小型蛋白质分子,因为OppA蛋白质有一个非常大的空腔,水分子原本填充在里面,一旦水分子被排出空腔,小蛋白质分子就可以填充进去,多余的空隙再由水分子填补结实。另一方面,位于蛋白质结构当中非常精确的位置的水分子可以跨越蛋白质表面以及其他分子表面的空隙,从而让蛋白质呈现特定的形状。画面上是血红素氧合酶分子具有催化活性的区域,可以看到这片区域有一张鲜明的小型网络,由紫色的水分子构成。这套网络传动并且协调了蛋白质链不同部分的运动,对于催化过程至关重要,因此实际上成为了蛋白质结构的一部分。如果水分子团被打乱了,那么酶分子就发挥不了原本的功用。
因此水分子可以被视为乐高砖块,能够以各种方式拼接在蛋白质上面。生化分子将这种乐高式的水分子团塑造成为了新的形状,这些形状未必总会出现在普通的水当中。尤其重要的是,狭窄的蛋白通道产生了许多种在生物学当中功能最多且最常见的水分子团。只要蛋白通路足够狭窄,一次只允许一个水分子通过,就可以形成一条由氢键链接而成的水分子长链或者说水线。之所以叫水线这个名字,不仅因为这样的结构又长又细,还因为水线就像铜线一样也可以传导电荷。但是铜线传导的是带负电的电子,水线传导的则是带正电的氢离子或者说质子。质子可以沿着水线迅速传播,因为只要水线上的一个水分子翻转一下方向,就可以让下一个水分子产生一个质子,换句话说并不需要一个实际的质子从水线一头传到另一头,水分子只需要进行翻转方向的接力就行,这使得质子在水线里传播的很快——这就好比牛顿摆,一侧的小球撞下去,另一侧的小球立刻就弹起来。正因为如此,氢离子在纯水当中传播得很快,因为纯水当中同样存在水线。
由酶参与的生化反应往往需要氢离子。多种酶都会使用水线将氢离子从外部溶液传输到酶的中心,在这里发生催化反应。比方说画面上展示的结构是细菌视紫红质,这种蛋白质存在于某种古老的单细胞光合作用生物体内,我们称之为古菌。细菌视紫红质本质上是一种由光驱动的质子泵,作用是将细胞内部的质子泵到细胞外部。光可以影响细菌视紫红质结构当中的绿色部分——这部分基本上相当于眼睛里的感光色素——一旦这部分遭到光的触发,就会释放一个质子,然后这个质子沿着氢键结合的水分子链传导出去,直到排出细胞之外。
另一类包含水通路的重要蛋白质名叫水孔蛋白。这种蛋白质的作用是调控流入与流出细胞的水流。水孔蛋白只允许水分子通过,不允许盐或者其他溶解的物质通过,换句话说水孔蛋白就是分子层面的过滤器。我们的肾脏正是倚靠水孔蛋白才能每天处理一百八十升液体。因为水孔蛋白让盐无法与水一起通过细胞膜,所以今天有人正在研究如何利用这种蛋白质来开发基于生物学原理的海水淡化系统。不过我们还可以利用同样的渗透原则来淡化海水,而不需要依赖娇贵的水孔蛋白。目前最看好的做法就是使用所谓的碳纳米管,也就是非常细的管子,直径只有若干纳米,由碳原子构成。这些管子的强度非常大,我们也非常擅长制造它们,只不过我们擅长制造的纳米管的直径区间很有限。只要将这些纳米管嵌入结实的薄膜,然后向海水施压,迫使其通过薄膜,那么渗过去的都是水,留下来的全是盐与其他污染物质。如果纳米管足够细,比方说直径小于一纳米,那么水分子就只能以单链结构通过。纳米管内壁疏水性极强,这意味着水分子通过的时候几乎不会受到阻力,因为内壁就像上过油一样。
更有甚者,盐无法通过纳米管。盐离子本身体积比水分子还小,照理说水分子过得去盐离子就该过得去,但它们就是过不去,因为单列通过纳米管的水分子把管道堵住了。盐溶解之后,离子包裹在水分子的笼子里,使得离子的体积变大,除非把所有水分子都去掉才能恢复本来的体积。这就好像妮可.基德曼想要穿过一扇门,我相信以她的身材走过绝大多数门户都轻松自如,但实际上却很困难。因为她身边有大量的狗仔队、粉丝和保镖,必须把所有其他人都去掉她才能走过去,而且驱散这些人肯定需要耗费大量能量。盐的情况也是一样。计算机模拟发现,直径8埃的纳米管就可以去掉水里所有的盐略。略微粗一点的纳米管——直径9埃——可以去掉95%的盐,这就足够将海水变成饮用水了。一平方米的纳米管膜理论上每天可以生成12亿升清水。与理论相差不远的膜已经造出来了。加州国立实验室已经做出了管道内径大约是三四十纳米的纳米管,将其嵌入陶瓷基体当中,可以过滤98%的盐离子。
就像纳米管一样,水孔蛋白让水通过而不让盐离子通过,但是它实际上更聪明一些,不仅能过滤盐离子,还能过滤氢离子,换句话说就是维持细胞内部的酸碱平衡。它解决这个问题的方式非常聪明。刚才我提到氢离子很容易沿着水线传递,那么水孔蛋白要怎样让水线通过同时又阻止氢离子?首先,水线通过的管壁上的氨基酸会调整水分子的方向,使其无法倒转;此外在管道出入口还有带负电的区域,排斥一切氢离子。这两种机制共同将氢离子排斥在外。自然界还有其他聪明的手段来控制水流通过水孔。画面上是镶嵌在某些细菌的细胞膜上面的另一种蛋白质,名叫MscS,功能是调控铁离子通过细胞膜,原理则是通过轻微机械形变改变通路的孔径宽度。这种通路通过细胞膜的机械变形开启与闭合。只要孔径缩小一点点,那么任何离子都过不去。研究人员正在思考能否让智能水过滤膜模拟这个过程,例如在纳米管内部添加化学基团充当开关与阀门。
我想现在我已经让大家理解了水为什么不仅仅是生命活动这幅精美挂毯的平淡背景。水本身也是一种生物分子,而且还是一种非常复杂的分子,可以参与很多生命过程,形成功能多样的宏观大分子,足以与蛋白质、核酸、细胞膜以及其他生物质相互作用。我认为这一理念提出新的问题:水对于生命是否不可或缺?Taylor & Francis出版社最近出版的新书《水与生命》就讨论了这个问题。如果我们认为所有足够复杂、可以被称为生命的化学过程都需要我们在蛋白质互动当中见到的各种特质,那就很难想象还有除了水以外的其他溶剂足以支持这种过程。水在生命活动当中展现的许多特质似乎都或多或少地取决于水分子本身的特殊性质,尤其是水分子惯于产生方向性的弱氢键这一点,因为这项特性使得水分子可以在三维空间反复重组,形成各种离散且可辨识的三维结构。因此尽管水在生命活动当中发挥的一部分作用换成其他溶剂也能完成,并不是水的专利,但是却很难想到哪种溶剂可以填补水的一切角色。
但是这并不等于说水对于生命不可或缺,一切生命必须基于水的前提是水的任何一种功能对于任何生命来说都不可或缺。我们完全可以想象甚至制造一套化学体系,使其可以进行某种形式的信息传递——信息传递正是达尔文式进化与遗传不可或缺的关键,有了信息传递才会有生命体的自我复制——而且确实存在可以在非水液体里做到这一点的化学体系。反过来说,以水为基础的生命也有不足之处。比如说水很容易发生化学反应,惯于裂解作为生命基石的长链大分子,例如蛋白质与碳水化合物。尽管NASA寻找可居住地外世界的原则是寻找水存在的迹象,但是天文生物学家却警告我们要小心这种水中心论,不要想当然地认为任何地方的生命都离不开水。我们完全可以想象生命出现在很多备选溶剂当中,前提是我们不能目光短浅地一口咬定生命必须由蛋白质和核酸构成。比方说木星上存在大量的氨,金星上存在大量硫酸,甲酰胺分子在宇宙当中非常常见,土卫六上存在大量液态甲烷。再冷门一点,超临界氢也可以充当溶剂,而且很可能在木星与其他气体大行星上大量存在。再进一步,就连气体也未必不能充当溶剂。弗莱德.霍尔(Fred Hoyle)在二十世纪六十年代创作的科幻小说《黑云》就探讨了气体生命的存在形式。
问题在于我们不知道怎样区分生命与非生命,甚至不知道这一疑问究竟是不是有意义的科学问题,但是通过设想非水基的地外生物学溶剂,今后我们再去回顾水在地球生命化学当中扮演的角色时难免想到,我们并不能理直气壮地设想生命的分子机器与承载这些分子的溶剂之间可以画出一条一清二楚的分界线。就水而言,分子机器与溶剂之间存在着双向的行为与信息交换,这样一来一切生物组件与环境之间的分界线就都被抹掉了。水是一种反应敏捷且高度交感的溶剂,远不仅仅是普通的溶剂,而且任何生命体系都需要这种交换,从而让分子信息可以传递到承载信息的分子之外。因此我们难免忍不住认为生命活动必须依靠水才能进行。
目前这些问题都是纯粹的假想,不过有两种方式可能突破假想的局限。首先,我们或许可以详细考察奇异的地外世界的表面。实际上我们已经这么做了。2004年,卡西尼-惠更斯号探测器飞临土星与土卫六,拍摄了许多惊人的土卫六地表图像,让我们看到了土卫六上的碳氢化合物大湖是什么样?NASA与ISA正在合作勘测土星与冰层覆盖的卫星,尤其是木卫二,因为我们相信在木卫二的冰盖下存在盐水构成的液态海洋。这项任务预定于2020年开展。此外我们已经收集了五百个地外行星的信息——也就是围绕其他恒星公转的行星——这使得我们在一定程度上理解了宇宙当中可能存在的世界有多么多种多样。其次,通过合成生命科技的进步,我们或许可以在实验室里人工构建生命一样的实体,可以存在于水之外的溶剂当中。仅就我个人而言,我其实并不敢肯定无水的生命究竟能不能行得通。但是我们至少可以确定,我们已知的所有生命形式都有理由感谢水的奇怪特质。谢谢大家。
- 相关回复 上下关系2
🙂261-Philip Ball:水为什么这么怪?
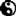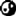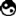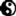
🙂有意思得很,谢谢你的分享 bluestarry1 字0 2023-06-03 01:33:46